Другой факт касается разведения гепардов в питомниках,
куда животные помещались для восстановления исчезающей популяции. Получая
питание вне естественной среды обитания из рук человека, животные демонстрировали
нарушение фертильности. Анализ диетических особенностей выявил избыточное
содержание соевых продуктов. После устранения «соевого перекоса» положение
выправилось.
Предположения о влиянии растений подтвердились в эксперименте
на животных. Позднее путем химического анализа было установлено, что
гормональная активность растений связана с наличием в их составе веществ,
близких по строению к эстрадиолу. В связи с этим гормоноактивные вещества
получили название фитоэстрогенов. Далее после появления сообщений о
том, что у сборщиц хмеля происходят нарушения менструального цикла вплоть
до временной аменореи, было выдвинуто предположение о похожем влиянии
и на человеческий организм.
Несколько позднее по времени проблема фитоэстрогенов
приобрела новый качественный оттенок. А именно, фитоэстрогены стали
рассматриваться, как средство профилактики гормонозависимых опухолей
– рака молочной железы и рака предстательной железы. Изменение научного
акцента произошло на волне всеобщего интереса к так называемому «японскому
феномену». Суть его заключается в том, что японки значительно реже,
нежели жительницы Европы и США, болеют раком молочной железы. После
того, как стало ясно, что решающее влияние оказывает стереотип питания,
были выделены его особенности. Они заключаются в употреблении большого
количества морепродуктов, водорослей, рыбы, а также овощей и особенно
сои. В последствии из сои были выделены флавоноиды генистеин и дайдзеин,
для которых на большом экспериментальном материале была доказана активность
по отношению к гормональной сфере. Был открыт факт связывания флавоноидов
с эстрогеновыми рецепторами, модулирующее влияние на эти рецепторы,
изменение под действием фитоэстрогенов активности ферментов, участвующих
в различных этапах синтеза половых стероидов и их печеночного и внепеченочного
превращения. Были проведены исследования, расширяющие сферу изучения
фитоэстрогенов как в сторону поиска других веществ в составе растений,
обладающих аналогичными свойствами, так и с точки зрения углубления
знаний в отношении их влияний на другие гормональные процессы. Был существенно
расширен круг онкологической и неонкологической патологии, при которой
ожидалось получение положительных эффектов от применения фитоэстрогенов.
В частности, изучена и обоснована роль фитоэстрогенов, как средства
профилактики и лечения постменопаузальных проблем – остеопороза и дистрофии
слизистых половых органов.
По мере углубления медицинских знаний о фитоэстрогенах,
были установлены также и негативные аспекты их влияния на организм.
Их нужно рассматривать в двух вариантах. Во-первых, это влияния, связанные
с поступлением фитоэстрогенов в организм в процессе специфической трудовой
деятельности или условий проживания. В данном контексте вещества принято
называть не фито-, а ксеноэстрогенами, подразумевая гораздо более широкую
группу веществ, обладающих проэстрогенной активностью, таких как продукты
промышленного загрязнения или медикаменты нерастительного происхождения.
А во-вторых, как это ни печально, аспекты неверного применения или чрезмерного
увлечения препаратами (чаще всего БАДами), содержащими фитоэстрогены.
Так, например, чрезмерное употребление БАДов, содержащих генистеин,
приводило к появлению гиперпластических процессов в эндометрии.
В настоящее время процесс углубления знаний о фитоэстрогенах
продолжается. Можно сказать, что в основном сформировался список растительных
веществ, обладающих свойствами фитоэстрогенов. В главных аспектах раскрыты
закономерности фармакологии данной группы веществ. Отработаны методики
определения количества и качества фитоэстрогенов в средах организма,
в растительном сырье и в готовых препаратах. Учитывая сложившуюся на
настоящий момент тенденцию, можно смело утверждать, что мы вправе рассчитывать
на получение еще более интересной информации в самые ближайшие годы.
Введение.
Видное место среди растений, применяемых для лечения
гормонозависимых опухолей (более 40% всех опухолей), занимают травы
с высоким содержанием фитоэстрогенов. Их лечебный эффект связывают со
способностью к избирательной блокаде эстрогеновых (и андрогеновых) рецепторов
на клетках опухолей, а также с изменением активности некоторых ферментных
систем, участвующих в синтезе и метаболизме гормонов.
Научные изыскания последних лет указывают на наличие
у фитоэстрогенов противоопухолевой активности, не имеющей отношения
к эндокринной регуляции. В частности, доказана способность флавоноидов
индуцировать и усиливать апоптоз, блокировать действие ростовых факторов
на клетку и моделировать иммунные реакции по отношению к злокачественной
клетке. В связи с этим противоопухолевый эффект фитоэстрогенов был зарегистрирован
в экспериментах с опухолями, не имеющими гормональной зависимости (толстый
кишечник, легкие, печень и др.).
Факт гормональной зависимости опухолей молочной железы,
эндометрия, предстательной железы в настоящее время ни у кого не вызывает
сомнений, так же как и объективность эффекта антигормональной терапии.
Однако вопрос о применении лекарственных растений в этой области нельзя
считать до конца определенным. С одной стороны, известно онкопрофилактирующее
действие фитоэстрогенов сои при раке молочной железы. С другой стороны,
логично было бы ожидать от растения с высокой гормональной активностью
провоцирующего усиление роста. В связи с этим нам показалось важным
определить показания и противопоказания к применению ФЭ-содержащих растений
в онкологии. Для этого решались следующие задачи:
1. Проводился анализ молекулярных эффектов фитоэстрогенов
2. Оценивалось распределение ФЭ по эстрогеноподобной
активности
3. Формировался список растений в соответствии с содержанием
ФЭ
Классификация фитоэстрогенов
Чаще всего встречается упрощенная классификация фитоэстрогенов
по химическому строению, в которой выделяются 3 группы:
1. Изофлавоны (генистеин, дайдзеин, апигенин, биоханин
и т.д.)
2. Лигнаны (энтеродиол, энтеролактон)
3. Куместаны (куместрол, веделолактон, пликадин)
Внутри первой группы химики выделяют порядка 15-17
подгрупп, что для медицинской практики пока не имеет особенного значения.
К изофлавонам относят генистеин, дайдзеин, апигенин, кверцетин и другие
вещества. Лигнаны представлены в основном двумя соединениями – энтеродиолом
и энтеролактоном. Куместаны рассматриваются на примере куместрола.
Имеются групповые различия в эффектах. Однако эффекты
изофлавонов существенно различаются, несмотря на принадлежность их к
одной химической группе. Таким образом, с точки зрения фармаколога далеко
не всегда имеет смысл классифицировать действующие вещества растений
по химическому признаку. Гораздо интереснее было бы видеть классификацию
по силе эстрогенного эффекта, по степени аффинитета к эстрогеновым рецепторам,
по преимущественному влиянию фитоэстрогенов на тот или иной подтип рецепторов,
и как следствие, на тот или иной гормонозависимый орган. Можно предложить
еще несколько признаков, по которым имело бы смысл классифицировать
фитоэстрогены для пользы клинической практики. К сожалению, количество
научной информации о фитоэстрогенах (хоть и довольно уже большое) еще
не перешло в новое качество, для которого характерны были бы подобные
классификации.
Фармакологические эффекты ФЭ, связанные
с эндокринной регуляцией
При изучении механизмов влияния фитоэстрогенов на
клетки гормонозависимых органов, а также системные аспекты гормональной
регуляции, чаще всего обсуждаются:
1. Селективная модуляция эстрогеновых (андрогеновых)
рецепторов
2. Изменение активности ароматазы и других ферментов
стероидогенеза
3. Оптимизация печеночного и кишечного метаболизма
гормонов
Селективная модуляция эстрогеновых
рецепторов.
Феномен эндокринной регуляции подразумевает существование
как минимум трех субъектов регуляции. Должен существовать регулирующий
сигнал, кроме того, регулятор, издающий этот сигнал и, наконец, регулируемый
субъект, воспринимающий сигнал. Так оно и есть: имеется эндокринная
железа (регулятор), вырабатывающая гормон (сигнал), который воспринимается
органом-мишенью (регулируемый субъект), вследствие чего функция последнего
изменяется.
Для того чтобы гормон воспринимался клеткой-мишенью
как совершенно конкретный сигнал, он должен иметь постоянную и неизменяемую
химическую структуру. А участок клеточной мембраны, отвечающий за восприятие
гормонального сигнала (рецептор), должен реагировать только на данный
конкретный гормон, и ни на что иное. Говорят, что гормон должен подходить
к своему рецептору, как ключ к замку.
Фитоэстрогены, не будучи полноценными с химической
точки зрения эстрогенами для человека, все же имеют структуру, весьма
близкую к эстрадиолу. Близкую ровно на столько, чтобы клеточный эстрогеновый
рецептор воспринял фитоэстроген, как «свой ключ», и наступило бы присоединение
молекулы фитоэстрогена к рецептору.
Процесс рецепции весьма сложен, и помимо соединения
гормона с рецептором, включает также конфирмационные изменения рецептора
под действием гормона, транспорт комплекса рецептор-гормон внутрь клетки
и внутрь клеточного ядра, и, наконец, активацию определенных участков
хромосомного аппарата. Таким образом, присоединение фитоэстрогена к
рецептору – это только полдела. Вторая половина зависит оттого, достанет
ли силы фитоэстрогену вызвать такие конфирмационные перестройки рецептора.
Понятно, что лучше всего подобное действие осуществит «родной» эстрадиол,
он вызовет самые сильный и бурный ответ клетки. Поставим его на одном
конце линейки веществ с эстрогенным действием, присвоив ему статус «абсолютного
плюса». Кто же тогда окажется на другом конце линейки? Это будет какое-то
гипотетическое вещество, которое с одной стороны обладает достаточным
сродством к рецептору, чтобы с ним связаться, а с другой стороны, имеет
особенности химической структуры, препятствующие активации рецептора.
Что же тогда получится? С одной стороны рецептор окажется блокированным,
и никто больше к нему присоединиться не сможет. С другой стороны, данный
связанный рецептор, выпавший из передачи сигнала, уже не в состоянии
подействовать на хромосомный аппарат. Клетка через это «ухо» эстрогеновый
сигнал уже «не услышит». Продолжая аналогию «ключ-замок», можно сказать
так: в замочную скважину можно вставить похожий ключ, но повернуть его
там и открыть дверь все равно не получится. В этом и заключается смысл
действия блокаторов эстрогеновых рецепторов, применяемых для лечения
рака молочной железы. Например, препарат тамоксифен. Присвоим ему статус
«абсолютный минус» и поставим на противоположный от эстрадиола край
линейки.
А что же фитоэстрогены, где они стоят на этой нашей
виртуальной линейки. Оказывается, что они распределяются по всей ее
длине. Часть из них близка к блокаторам, другая к эстрадиолу, а кто-то
расположен где-то посередине. Получается, что при помощи фитоэстрогенов
можно добиваться самого разнопланового эффекта на эстрогеновый рецептор
– и блокирования, и наоборот стимуляции. Поэтому фитоэстрогены принято
называть SERM – от английского Selective Estrogen-Receptor Modulator,
то есть селективный модулятор эстрогеновых рецепторов. В настоящее время
существует фармакологическая группа синтетических препаратов, первый
из которых, тамоксифен, уже упоминался выше. Он давно используется для
лечения рака молочной железы. Ему на смену уже подошел более совершенный
ралоксифен (Эвиста). А в научной разработке имеются еще несколько.
Как уже говорилось выше, поиск антиэстрогенных препаратов
был продиктован необходимостью лечить рак молочной железы. Начиная применять
тамоксифен, врачи ожидали появление побочных эффектов со стороны других
органов, лишенных действия эстрогенов. Однако не вполне ожидаемым побочным
эффектом явилась увеличивающаяся частота гиперпластических процессов
в эндометрии, полипов и даже карцином. Данная ситуация не могла не заинтересовать
исследователей, и в ходе изучения оказалось, что существует как минимум
две субпопуляции эстрогеновых рецепторов – альфа и бета – неодинаково
реагирующих на ксеноэстрогены. В частности, ЭР-альфа блокируется тамоксифеном,
а ЭР-бета, напротив, стимулируется. Кроме того, оказалось, что плотность
эстрогеновых рецепторов разных субпопуляций в разных эстрогенозависимых
органах различается. Так, вполне объяснимо в молочной железе больше
альфа-рецепторов, а в эндометрии – бета. Какое значение это имеет в
контексте нашего разговора о фитоэстрогенах? На волне «соевого бума»
появилось много БАДов, содержащих экстракт соевых бобов и даже очищенные
флавоноиды генистеин и дайдзеин. Эти препараты стали активно применяться
женщинами, как по назначению врача, так и без него. Со временем стали
появляться сообщения о более частом выявлении гиперплазии эндометрия
у женщин, слишком активно употреблявших БАД из сои. Последовавшие за
этим эксперименты доказали стимулирующее влияние генистеина и дайдзеина
на ЭР-бета.
Наибольшей эстрогеноподобной активностью обладают хмель,
солодка, клевер, люцерна, душица. Так, например, эстрогенную активность
хмеля, активность которого, по-видимому, связана в первую очередь с
изоксантохумолом и пренилнарингенином, исследовали методом Аллена-Дойзи
на кастрированных мышах и инфантильных крысах. У крыс экстракт хмеля
вызывает появление эструса или проэструса. Активность 1 кг сухого хмеля,
экстрагированного водой, составила 1000 мышиных единиц. Наиболее активной
оказалась фенольная фракция хмеля, обладающая эстрогенной активностью,
равной 25 000 мышиных единиц (на 1 кг сухого хмеля). Фенольная фракция
в дозе 10-30 мг на мышь и 20-40 мг на крысу вызывала эстрогенный эффект
у 80% животных. В гениталиях обнаруживались изменения, характерные для
эстрогенного действия: влагалище было заполнено студенистой жидкостью,
рога матки утолщены, масса гениталий подопытных животных была в 2,7
раза выше, чем у контрольных животных.
По мнению А. Ф. Гаммерман (1976) солодка имеет самую
высокую эстрогенную активность из кормовых растений. Установлено, что
солодка обладает эстрогенной активностью, превышающей 10 000 ME в 1
кг сухой массы, тогда как у люцерны и клевера она не выше 1300 ME (Д.
А. Муравьева, 1978). Эстрогенная активность солодки голой связана с
наличием в ней эстриола (И. А. Муравьев и соавт., 1972). В то же самое
время известно, что солодка содержит формононетин, ответственный за
эстрогеноподобное действие клевера, люцерны и софоры желтеющей.
Таблица №1. Сводная таблица
растений, содержащих фитоэстрогены.
| Фитоэстроген |
Растения |
Эстрогенная активность |
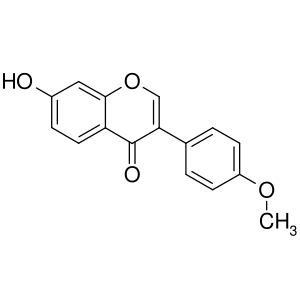
Генистеин
|
Соя, галега, клевер, жарновец метельчатый |
Высокая, избирательная |
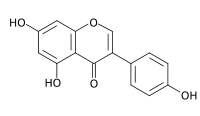 |
| дайдзеин |
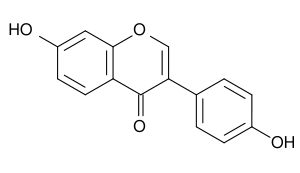 |
| Изоксантохумол |
Хмель, клевер красный |
Высокая |
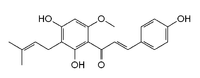 |
| 8-пренилнарингенин |
| Биоханин-А |
Клевер, люцерна |
Высокая |
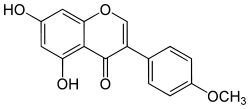 |
| Биоханин-В (формононетин) |
Солодка, софора желтеющая, люцерна |
Высокая |
 |
| Куместрол |
Клевер, люцерна, подсолнечник, пророщенные
зерна злаковых, пуэрария |
умеренная |
 |
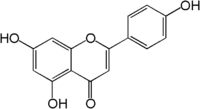
| Апигенин |
Ромашка, шлемник, осот, авран, береза,
бессмертник, валериана, волдырник, гладыш, горечавник, мытник, подорожник,
петрушка, солодка, чабрец, тысячелистник, чистец, шалфей |
отсутствует |
 |
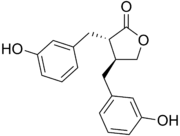
| Энтеродиол, энтеролактон |
Лен, пшеница, рожь, орехи, рис, вишня,
яблоко, морковь |
слабая |
 |
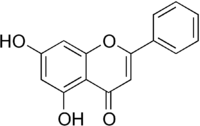
| Хризин |
Пикульник, тополь, шлемник, |
слабая |
 |
| Кемпферол |
Астрагал шерстистоцветковый, белокопытник,
береза, бересклет, бессмертник, бодяк, борщевик, брусника, валериана,
василек, василистник, володушка, галега, герани, горцы, горичник,
зверобой, каштан, клевер, клоповник, княжик, колокольчик и т.д.
Не менее 183 растений |
отсутствует |
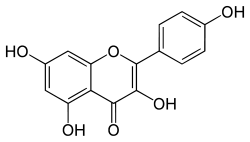 |
| Нарингенин |
Бессмертник, василек, ива, щавель конский,
яблоня |
слабая |
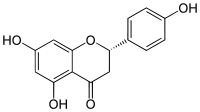 |
| Ресвератрол |
виноград |
слабая |
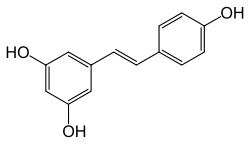 |
Таким образом, мы имеем все основания для введения ограничений
по использованию солодки, хмеля, клевера и люцерны у больных раком молочной
железы, по крайней мере, у тем, что имеют положительный иммуногистохимический
рецепторный профиль. И напротив, данные растения могут быть рекомендованы
для применения в комплексной терапии больных раком предстательной железы.
Влияние на ферменты стероидогенеза.
Существующие в настоящее время подходы к антигормональной терапии в
онкологии, в частности, рака молочной железы, ориентированы на препараты
двух групп. Об антиэстрогеновых препаратах мы уже сказали. Вторая группа
– ингибиторы ароматазы. Препараты данной группы уже зарекомендовали
себя с хорошей стороны. В связи с этим вполне объясним научный интерес
к вопросу о том, а как же действуют на ферменты стероидогенеза фитоэстрогены.
Был проведен целый ряд научных изысканий, результаты которых мы суммировали
в двух последующих таблицах, разделив эффекты фитоэстрогенов по сфере
испытания – эндометрий и молочная железа. На рисунке №1 приводится схема
синтеза половых стероидов, соотнося с которой наши таблицы, можно представить
точки приложения эффектов фитоэстрогенов. Наибольший интерес представляют
растительные вещества, способные по аналогии с соответствующими медикаментами
блокировать ароматазу (CYP19). В культуре клеток молочной железы таковыми
являются ресвератрол, энтеролактоны и в меньшей степени генистеин. В
эндометрии по антиароматазному действию лидируют апигенин, хризин, нарингенин
и кемпферол. У генистеина обнаружен стимулирующий эффект по отношению
к ароматазе в эндометрии.
Таблица №2. Изменение активности ферментов стероидогенеза
в эндометрии под влиянием фитоэстрогенов.
| |
ароматаза |
3-бета-HSD |
17-бета-HSD |
| генистеин |
стимулирует |
подавляет |
подавляет |
| дайдзеин |
0 |
подавляет |
подавляет |
| биоханин A |
0 |
подавляет + |
0 |
| апигенин |
подавляет + |
|
|
| энтеродиол |
0 |
0 |
0 |
| энтеролактон |
0 |
0 |
0 |
| зеараленон |
подавляет + |
|
подавляет + |
| нарингенин |
подавляет |
|
|
| хризин |
подавляет |
|
|
| кверцетин |
0 |
0 |
подавляет |
| куместрол |
|
подавляет |
|
| кемпферол |
подавляет |
|
|
Таблица №3. Изменение активности ферментов
стероидогенеза в клетках молочной железы под влиянием фитоэстрогенов.
| |
ароматаза |
17-бета-HSD |
| генистеин |
подавляет |
усиливает |
| энтеродиол |
подавляет + |
подавляет + |
| энтеролактон |
подавляет + |
подавляет + |
| ресвератрол |
подавляет + |
|
Таким образом, наличие антиароматазного эффекта у многих
фитоэстрогенов, пусть и выраженного в разной степени, позволяет позиционировать
ФЭ-содержащие растения в качестве средства многофакторного влияния на
гормонозависимые виды опухолей молочной железы и эндометрия. В последней
случае применение, растений, содержащих генистеин, является нежелательным.
«Негормональные» механизмы действия
фитоэстрогенов - индукция апоптоза.
Растительные вещества, названные фитоэстрогенами по
причине их способности оказывать эстрогеноподобное влияние, отнюдь не
ограничивают свой эффект гормональным рецептором и ферментами стероидогенеза.
Не удивительно, что у многих из них был обнаружен цитостатический эффект
по отношению как к гормононезависимым видам рака молочной железы и предстательной
железы, так и к опухолям, изначально не относящимся к гормонозависимой
сфере – рак легких, рак толстого кишечника, рак щитовидной железы, рак
кожи и т.п. На клеточном уровне данное воздействие выражается в усилении
апоптоза, в развитии которого были экспериментально распознаны следующие
механизмы:
• Подавление экспрессии генов, отвечающих за синтез
антиапоптогенных белков Bcl-2 и возрастание коэффициента Bax/Bcl2
• Усиление конфирмационных изменений Bax с последующей
транслокацией его из цитозоля в митохондрии, приводящие к уменьшению
мембранного потенциала и высвобождением цитохрома С. Появление цитохрома
С в цитозоле активирует капсазу-9 и капсазу-3.
• Подавление экспрессии гена, отвечающего за синтез
белка – ингибитора апоптоза (IAPs) .
• Фосфорилирование митоген-активированной протеинкиназы
(МАРК), возрастание внутриклеточного Са++, приводящие к активному образованию
агрессивных соединений кислорода.
• Образование ДНК-комплексов с топоизомеразой-I и топоизомеразой-II,
запускающее клеточную гибель.
• Усиление апоптоза, возникающего под действием химиопрепаратов
(апигенин+5ФУ)
• Блокирование клеточного цикла злокачественных клеток
в S- и G2/M-фазах.
Можно с уверенностью утверждать, что для клинической
практики, когда формирование противоопухолевого эффекта осуществляется
благодаря совокупности многочисленных терапевтических направлений, не
меньшую важность имеют доказанные антиоксидантный, антигипоксантный,
противовоспалительный, иммуномодулирующий, вазоактивный и прочие эффекты
фитоэстрогенов.
Показания к применению фитоэстроген-содержащих
растений.
• Рак молочной железы
• Рак эндометрия
• Рак яичников
• Рак предстательной железы
• Мастопатия
• Доброкачественная гиперплазия предстательной железы
• Гиперплазия и полипы эндометрия
Выводы
Исходя из всего сказанного выше, очевидным становится,
что при раке молочной железы и эндометрия противопоказано применение
растений, содержащих ФЭ с высокой эстрогеноподобной активностью, например,
хмеля и солодки. Однако творческий подход к выбору трав требует различать
эффект мощных проэстрогеновых растений, содержащих генистеин и дайдзеин
(галега, дрок, соя и т.п.), которые неодинаково действуют на подгруппы
рецепторов к эстрадиолу. Можно получить хороший лечебный эффект при
опухолях молочной железы, в тоже самое время создать угрозу прогрессирования
гиперпластических процессов в эндометрии.
Напротив, рак предстательной железы, как и доброкачественная
гиперплазия предстательной железы, напротив, требуют назначения растений
с максимальной эстрогенопозитивной активностью.
Растения, содержащие энтеролактоны, в силу довольно
слабого эстрогенного действия, скорее всего, должны оцениваться как
слабые SERM`ы, подходят прежде всего для профилактики онкопатологии.
В силу того, что эти вещества в основном содержатся в злаковых культурах,
ягодах, орехах, здесь нужно говорить не столько о фитотерапии, сколько
лечебном питании.
Лекарственные растения, преимущественно содержащие
флавоноиды с низкой эстрогенопозитивной активностью, в особенности апигенин,
хризин, лютеолин (шлемник байкальский, осот розовый, ромашка аптечная
и т.п.), могут применяться при опухолях с любой степенью гормональной
зависимости.
Литература
1. Берштейн Л.М. Гормональный канцерогенез. – СПб.:
«Наука», 2000. – 200 с.
2. Берштейн Л.М. Онкоэндокринология: Традиции, современность, перспективы.
– СПб.: «Наука», 2004. – 343 с.
3. Бохман Я.В. Лекции по онкогинекологии. – М.: ООО «Медицинское информационное
агенство», 2007. – 304 с.
4. Гарин А.М. Эндокринная терапия и гормонозависимые опухоли. – М. –
Тверь: ООО «Издательство «Триада»», 2005. – 240 с.
5. Кит С. М., Турчин И. С. Лекарственные растения в эндокринологии –
К.: Здоров'я, 1986. — 80 с.
6. Манухин И.Б., Тумилович Л.Г., Геворкян М.А. Клинические лекции по
гинекологической эндокринологии, – М.: ООО «Медицинское информационное
агентство», 2001.
7. Практическая маммология. Под ред. М.И. Давыдова и В.П. Летягина –
М.: Практическая медицина, 2007. – с. 29 – 52.
8. Сидоренко Л.И. Гормонотерапия предрака и рака молочной желез. – Ленинград.:
Медицина. – 1986.
9. Limer JL, Parkes AT, Speirs V. Differential response to phytoestrogens
in endocrine sensitive and resistant breast cancer cells in vitro.Int
J Cancer. 2006 Feb 27;
10. Power KA, Ward WE, Chen JM, Saarinen NM, Thompson LU. Genistein
alone and in combination with the mammalian lignans enterolactone and
enterodiol induce estrogenic effects on bone and uterus in a postmenopausal
breast cancer mouse model. Bone. 2006 Feb 7;
11. Caetano B, Le Corre L, Chalabi N, Delort L, Bignon YJ, Bernard-Gallon
DJ. Soya phytonutrients act on a panel of genes implicated with BRCA1
and BRCA2 oncosuppressors in human breast cell lines. Br J Nutr. 2006
Feb;95(2):406-13.
12. Buterin T, Koch C, Naegeli H. Convergent transcriptional profiles
induced by endogenous estrogen and distinct xenoestrogens in breast
cancer cells. Carcinogenesis. 2006 Feb 10;
13. Nagel G, Mack U, von Fournier D, Linseisen J. Dietary phytoestrogen
intake and mammographic density -- results of a pilot study. Eur J Med
Res. 2005 Sep 12;10(9):389-94
14. Saarinen NM, Power K, Chen J, Thompson LU. Flaxseed attenuates the
tumor growth stimulating effect of soy protein in ovariectomized athymic
mice with MCF-7 human breast cancer xenografts. Int J Cancer. 2006 Mar
23;
15. Hertrampf T, Schmidt S, Seibel J, Laudenbach-Leschowsky U, Degen
GH, Diel P. Effects of genistein on the mammary gland proliferation
of adult ovariectomised wistar rats. Planta Med. 2006 Mar;72(4):304-10.
16. Gikas PD, Mokbel K. Phytoestrogens and the risk of breast cancer:
a review of the literature. Int J Fertil Womens Med. 2005 Nov-Dec;50(6):250-8.
17. Seo HS, Denardo DG, Jacquot Y, Laios I, Vidal DS, Zambrana CR, Leclercq
G, Brown PH. Stimulatory effect of genistein and apigenin on the growth
of breast cancer cells correlates with their ability to activate ER
alpha. Breast Cancer Res Treat. 2006 Mar 16;
18. Gallo D, Ferlini C, Fabrizi M, Prislei S, Scambia G. Lack of stimulatory
activity of a phytoestrogen-containing soy extract on the growth of
breast cancer tumors in mice. Carcinogenesis. 2006 Jan 7;
19. Rice S, Mason HD, Whitehead SA. Phytoestrogens and their low dose
combinations inhibit mRNA expression and activity of aromatase in human
granulosa-luteal cells., J Steroid Biochem Mol Biol. 2006 Nov;101(4-5):216-25.
Epub 2006 Sep 11.
20. Lacey M, Bohday J, Fonseka SM, Ullah AI, Whitehead SA. Dose-response
effects of phytoestrogens on the activity and expression of 3beta-hydroxysteroid
dehydrogenase and aromatase in human granulosa-luteal cells. J Steroid
Biochem Mol Biol. 2005 Aug;96(3-4):279-86
21. Whitehead SA, Lacey M. Phytoestrogens inhibit aromatase but not
17beta-hydroxysteroid dehydrogenase (HSD) type 1 in human granulosa-luteal
cells: evidence for FSH induction of 17beta-HSD. Hum Reprod. 2003 Mar;18(3):487-94
22. Whitehead SA, Cross JE, Burden C, Lacey M. Acute and chronic effects
of genistein, tyrphostin and lavendustin A on steroid synthesis in luteinized
human granulosa cells. Hum Reprod. 2002 Mar;17(3):589-94.
23. Edmunds KM, Holloway AC, Crankshaw DJ, Agarwal SK, Foster WG. The
effects of dietary phytoestrogens on aromatase activity in human endometrial
stromal cells. Reprod Nutr Dev. 2005 Nov-Dec;45(6):709-20.
24. Blomquist CH, Lima PH, Hotchkiss JR. Inhibition of 3alpha-hydroxysteroid
dehydrogenase (3alpha-HSD) activity of human lung microsomes by genistein,
daidzein, coumestrol and C(18)-, C(19)- and C(21)-hydroxysteroids and
ketosteroids. Steroids. 2005 Jul;70(8):507-14. Epub 2005 Mar 19.
25. Brooks JD, Thompson LU. Mammalian lignans and genistein decrease
the activities of aromatase and 17beta-hydroxysteroid dehydrogenase
in MCF-7 cells. J Steroid Biochem Mol Biol. 2005 Apr;94(5):461-7. Epub
2005 Mar 16.
26. Chen S, Zhang F, Sherman MA, Kijima I, Cho M, Yuan YC, Toma Y, Osawa
Y, Zhou D, Eng ET. Structure-function studies of aromatase and its inhibitors:
a progress report. J Steroid Biochem Mol Biol. 2003 Sep;86(3-5):231-7.
27. Papiez M, Gancarczyk M, Bilinska B. The compounds from the hollyhock
extract (Althaea rosea Cav. var. nigra) affect the aromatization in
rat testicular cells in vivo and in vitro. Folia Histochem Cytobiol.
2002;40(4):353-9
28. Saarinen NM, Huovinen R, Warri A, Makela SI, Valentin-Blasini L,
Sjoholm R, Ammala J, Lehtila R, Eckerman C, Collan YU, Santti RS. Enterolactone
inhibits the growth of 7,12-dimethylbenz(a)anthracene-induced mammary
carcinomas in the rat. Mol Cancer Ther. 2002 Aug;1(10):869-76
29. Brueggemeier RW, Gu X, Mobley JA, Joomprabutra S, Bhat AS, Whetstone
JL.
30. Yiwei Li, Mingxin Che, Sunita Bhagat, Kerrie-Lynn Ellis, Omer Kucuk,
Daniel R. Doerge, Judith Abrams, Michael L. Cher, and Fazlul H. Sarkar
Regulation of Gene Expression and Inhibition of Experimental Prostate
Cancer Bone Metastasis by Dietary Genistein. Neoplasia. 2004 July; 6(4):
354–363.
31. Delmulle L, Bellahcene A, Dhooge W, Comhaire F, Roelens F, Huvaere
K, Heyerick A, Castronovo V, De Keukeleire D. Anti-proliferative properties
of prenylated flavonoids from hops (Humulus lupulus L.) in human prostate
cancer cell lines. Phytomedicine. 2006 Nov;13(9-10):732-4. Epub 2006
May 5
32. Dalais FS, Meliala A, Wattanapenpaiboon N, Frydenberg M, Suter DA,
Thomson WK, Wahlqvist ML. Effects of a diet rich in phytoestrogens on
prostate-specific antigen and sex hormones in men diagnosed with prostate
cancer. Urology. 2004 Sep;64(3):510-5
33. Raschke M, Wahala K, Pool-Zobel BL. Reduced isoflavone metabolites
formed by the human gut microflora suppress growth but do not affect
DNA integrity of human prostate cancer cells. Br J Nutr. 2006 Sep;96(3):426-34
34. Cao F, Jin TY, Zhou YF Inhibitory effect of isoflavones on prostate
cancer cells and PTEN gene. Biomed Environ Sci. 2006 Feb;19(1):35-41
35. Staar S, Richter DU, Makovitzky J, Briese V, Bergemann C. Stimulation
of endometrial glandular cells with genistein and daidzein and their
effects on ERalpha- and ERbeta-mRNA and protein expresion. Anticancer
Res. 2005 May-Jun;25(3A):1713-8
36. Rannikko A, Petas A, Rannikko S, Adlercreutz H. Plasma and prostate
phytoestrogen concentrations in prostate cancer patients after oral
phytoestogen supplementation. Prostate. 2006 Jan 1;66(1):82-7
37. Rannikko A, Petas A, Raivio T, Janne OA, Rannikko S, Adlercreutz
H. The effects of short-term oral phytoestrogen supplementation on the
hypothalamic-pituitary-testicular axis in prostate cancer patients.
Prostate. 2006 Jul 1;66(10):1086-91
38. Rice L, Samedi VG, Medrano TA, Sweeney CA, Baker HV, Stenstrom A,
Furman J, Shiverick KT. Mechanisms of the growth inhibitory effects
of the isoflavonoid biochanin A on LNCaP cells and xenografts. Prostate.
2002 Aug 1;52(3):201-12.
39. Schabath MB, Hernandez LM, Wu X, Pillow PC, Spitz MR. Dietary phytoestrogens
and lung cancer risk. JAMA. 2005 Sep 28;294(12):1493-504
40. Sun AS, Yeh HC, Wang LH, Huang YP, Maeda H, Pivazyan A, Hsu C, Lewis
ER, Bruckner W, Fasy TM. Pilot study of a specific dietary supplement
in tumor-bearing mice and in stage IIIB and IV non-small cell lung cancer
patients. Nutr Cancer. 2001;39(1):85-95
41. Li D, Yee JA, McGuire MH, Murphy PA, Yan L. Soybean isoflavones
reduce experimental metastasis in mice. J Nutr. 1999 May;129(5):1075-8
42. Menon LG, Kuttan R, Nair MG, Chang YC, Kuttan G. Effect of isoflavones
genistein and daidzein in the inhibition of lung metastasis in mice
induced by B16F-10 melanoma cells. Nutr Cancer. 1998;30(1):74-7
43. Lee YS, Kim TH, Cho KJ, Jang JJ. Inhibitory effects of biochanin
A on benzo(a)pyrene induced carcinogenesis in mice. In Vivo. 1992 May-Jun;6(3):283-6
44. Lee YS, Seo JS, Chung HT, Jang JJ. Inhibitory effects of biochanin
A on mouse lung tumor induced by benzo(a)pyrene. J Korean Med Sci. 1991
Dec;6(4):325-8
45. Gilad LA, Tirosh O, Schwartz B. Phytoestrogens regulate transcription
and translation of vitamin D receptor in colon cancer cells. J Endocrinol.
2006 Nov;191(2):387-98
46. Sung MK, Lautens M, Thompson LU. Mammalian lignans inhibit the growth
of estrogen-independent human colon tumor cells. Anticancer Res. 1998
May-Jun;18(3A):1405-8.
47. Sorensen IK, Kristiansen E, Mortensen A, Nicolaisen GM, Wijnands
JA, van Kranen HJ, van Kreijl CF. The effect of soy isoflavones on the
development of intestinal neoplasia in ApcMin mouse. Cancer Lett. 1998
Aug 14;130(1-2):217-25
48. Yu J, Cheng Y, Xie L, Zhang R. Effects of genistein and daidzein
on membrane characteristics of HCT cells. Nutr Cancer. 1999;33(1):100-4
49. Davies MJ, Bowey EA, Adlercreutz H, Rowland IR, Rumsby PC. Effects
of soy or rye supplementation of high-fat diets on colon tumour development
in azoxymethane-treated rats. Carcinogenesis. 1999 Jun;20(6):927-31
50. Kallay E, Adlercreutz H, Farhan H, Lechner D, Bajna E, Gerdenitsch
W, Campbell M, Cross HS. Phytoestrogens regulate vitamin D metabolism
in the mouse colon: relevance for colon tumor prevention and therapy.
J Nutr. 2002 Nov;132(11 Suppl):3490S-3493S
51. Lechner D, Cross HS. Phytoestrogens and 17beta-estradiol influence
vitamin D metabolism and receptor expression-relevance for colon cancer
prevention. Recent Results Cancer Res. 2003;164:379-91
52. Cross HS, Kallay E, Farhan H, Weiland T, Manhardt T. Regulation
of extrarenal vitamin D metabolism as a tool for colon and prostate
cancer prevention. Recent Results Cancer Res. 2003;164:413-25
53. Guo JM, Xiao BX, Liu DH, Grant M, Zhang S, Lai YF, Guo YB, Liu Q.
Biphasic effect of daidzein on cell growth of human colon cancer cells.
Food Chem Toxicol. 2004 Oct;42(10):1641-6
54. Linsalata M, Russo F, Notarnicola M, Guerra V, Cavallini A, Clemente
C, Messa C. Effects of genistein on the polyamine metabolism and cell
growth in DLD-1 human colon cancer cells. Nutr Cancer. 2005;52(1):84-93
55. Danbara N, Yuri T, Tsujita-Kyutoku M, Tsukamoto R, Uehara N, Tsubura
A. Enterolactone induces apoptosis and inhibits growth of Colo 201 human
colon cancer cells both in vitro and in vivo. Anticancer Res. 2005 May-Jun;25(3B):2269-76
56. Lechner D, Bajna E, Adlercreutz H, Cross HS. Genistein and 17beta-estradiol,
but not equol, regulate vitamin D synthesis in human colon and breast
cancer cells. Anticancer Res. 2006 Jul-Aug;26(4A):2597-603
57. Zeleniuch-Jacquotte A, Lundin E, Micheli A, Koenig KL, Lenner P,
Muti P, Shore RE, Johansson I, Krogh V, Lukanova A, Stattin P, Afanasyeva
Y, Rinaldi S, Arslan AA, Kaaks R, Berrino F, Hallmans G, Toniolo P,
Adlercreutz H. Circulating enterolactone and risk of endometrial cancer.
Int J Cancer. 2006 Nov 15;119(10):2376-81
58. Pfeiffer E, Graf E, Gerstner S, Metzler M. Stimulation of estradiol
glucuronidation: a protective mechanism against estradiol-mediated carcinogenesis?
Mol Nutr Food Res. 2006 Apr;50(4-5):385-9




